

出品:科普中国
作者:黎茵(中山大学生命科学学院)
监制:中国科普博览
生物体是在重力的作用下进化而来的,植物能够感知重力的方向,从而调节地上部分和根系的形态和生长状态。
根系具有向地性(Gravitropism,又称为向重力性),是植物适应陆地环境最重要的过程之一。植物的地上部分向上生长,而根则一个劲儿往土里钻。根系向着重力方向生长,即便不是垂直放置甚至在被放平的状态下,都可以向着重力方向弯曲生长。

油菜幼苗向重力方向弯转(与刚开始平放时的照片相比较)
(图片来源:参考文献[1])
那这一切究竟是怎么发生的呢?
平衡细胞:我的作用是感受重力
根系表现出的向地性是一个非常复杂的生理过程。目前已经知道的是,根系对重力方向的感知,是来自于根冠中柱细胞中被称为“平衡细胞”的特化细胞。这些细胞内含有被称为“平衡石”(statolith)的淀粉体,淀粉体在根尖的敏感区域的沉积或者移动,起着平衡和引导生长的作用。平衡石可以在细胞中自由悬浮,当植物处于竖直状态时,平衡石会沉积在根尖的底部。
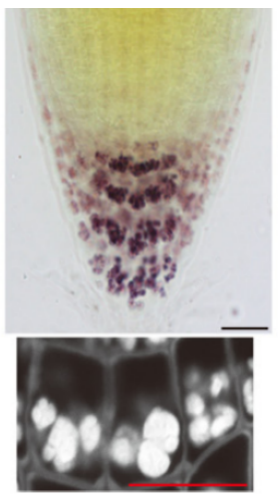
萌发7天的垂直生长的拟南芥幼苗根尖,在碘液染色和荧光染色下,细胞中淀粉体的分布情况
(图片来源:参考文献[7])
当植物根系发生倾斜时,平衡石由于重力沉积向细胞底部移动,其位置的改变会产生信号,传递到植物细胞,使植物能够重新调整生长方向,保持垂直生长。
因此,作为平衡石的淀粉体在根系的生长和重力感知中起着关键作用,并以此适应环境的变化。平衡细胞中淀粉体的移动会导致植物激素生长素定向流动的信号改变。生长素在介导根系向重力性中起着核心作用,并且能够促进重力感知,但具体的分子机制大部分尚未知晓。
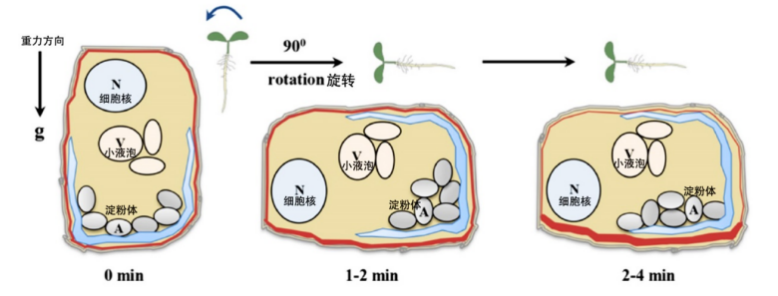
根冠细胞中的重力感应示意图。设定根垂直生长状态的起始时间为0分钟,旋转90度重新定向幼苗并开始计时,图示为重力刺激产生并启动重力感应的时间过程。
(图片来源:参考文献[5])
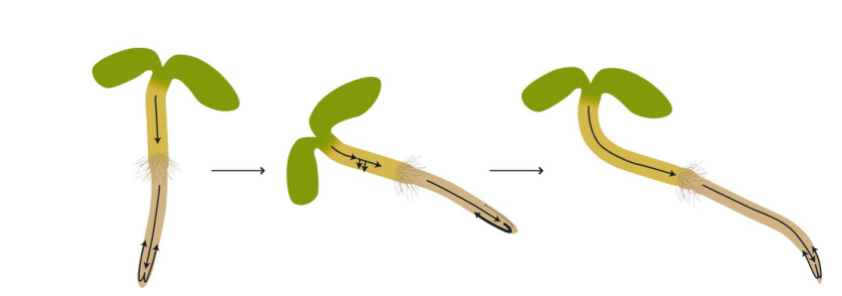
双子叶植物幼苗生长素的极性转运和向重力性。生长素运输的方向由箭头指示,其粗细表示在相应流中运输的生长素的量。
(图片来源:参考文献[6])
淀粉体:我不是力的搬运工
长期以来,重力传感机制一直被推测认为是淀粉体将力传递到细胞内结构的机械传感过程,这就是机械转导模型,但相关的分子机制并未得到过证明。
而之后的研究则提出另一种推测,也就是位置传感器模型,认为淀粉体接近或者触碰质膜,在接触位点诱导了细胞中的局部生长素流动,这些局部流量的总和决定了生长素总流量的方向。
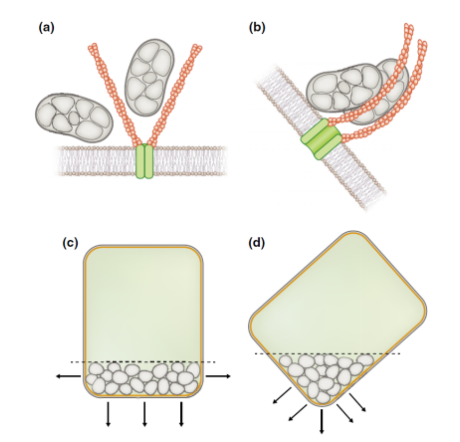
淀粉体重力信号转导的两种模型:机械转导模型(a,b),在稳定状态下(a),淀粉体不会通过挤压肌动蛋白丝来触发信号传导;一旦淀粉体响应重力刺激而移动(b),它们就会在肌动蛋白丝上提供机械张力以激活信号通道。位置传感器模型(c,d),淀粉质体和质膜之间的接近或接触位点诱导了细胞中的局部生长素流动,如箭头所示。
(图片来源:参考文献[2])
此前,科学家在水稻中发现一种LAZY1(懒惰)蛋白基因,这个基因突变后,水稻表现为茎匍匐生长的表型,就像爬不起来的懒虫。研究证实LAZY1蛋白与水稻茎的重力方向感知密切相关。随后的研究发现,植物细胞中淀粉体沉积信号的转导依赖于这种蛋白的家族成员(统称为LAZY1-LIKE蛋白,缩写LZY),但LZY蛋白与生长素转运的重定向过程之间的联系依然未能完全解析清楚。
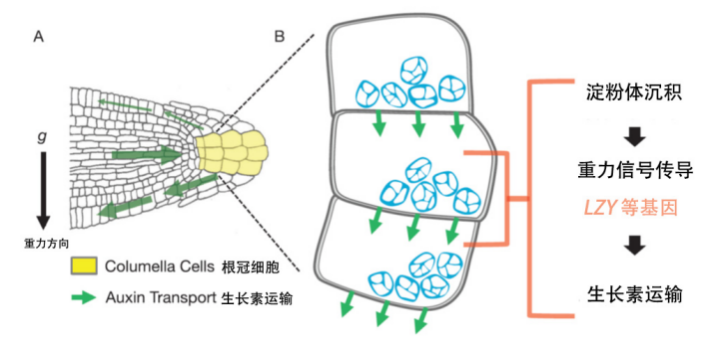
重新定向时初级根尖中生长素极性转运及根冠细胞重力信号传导过程示意图。
(图片来源:参考文献[3])
2003年8月在科学杂志(Science)发表的一项研究结果揭开了这个谜题,证明了LZY蛋白家族参与了植物根系淀粉体及相关质膜的重力信号转导,从而导致了根据重力方向的极性定位。
研究发现,拟南芥中所有的LZY蛋白的氨基酸序列中都有两个以上的高碱性疏水结构域,这些结构域通常与膜结合相关。如果将这些蛋白的基因进行突变,突变体植物的初生根就无法感应重力的方向,产生了不是往地下钻而是翘着向上长的奇特现象。通过转基因恢复LZY蛋白的表达,植物就能恢复正常,证明LZY蛋白在质膜上的定位是其在重力信号中发挥功能所必需的。
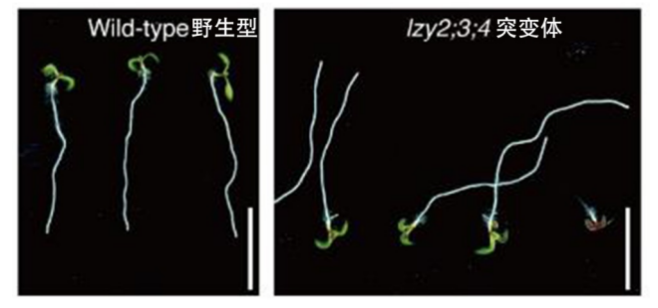
萌发5天的野生型和LZY突变体拟南芥幼苗
(图片来源:参考文献[4])
实验观察发现,在重力刺激作用下,淀粉体发生了移动,同时LZY蛋白也随重力方向转移到新的质膜位置,产生新的极性分布。而在一种淀粉缺失的突变体植物中,淀粉体基本无法向重力方向沉积,这时LZY蛋白也不能在新的质膜上产生极性。
科学家由此证明,LZY蛋白在质膜上的极化是淀粉体沉积导致的。通过对淀粉体的移动进行光学镊子操作,光学镊子使淀粉体发生位置移动(光学镊子是获得2018年诺贝尔物理学奖的一项技术,利用激光产生的辐射压对微小颗粒物质的移动进行无接触操纵)。
观察发现,LZY蛋白可以与淀粉体结合,并且会在淀粉体集中部位附近的质膜上聚集,这一过程并不需要重力的参与。由此可见,LZY蛋白的分布由淀粉体的位置决定,而不是由重力决定。
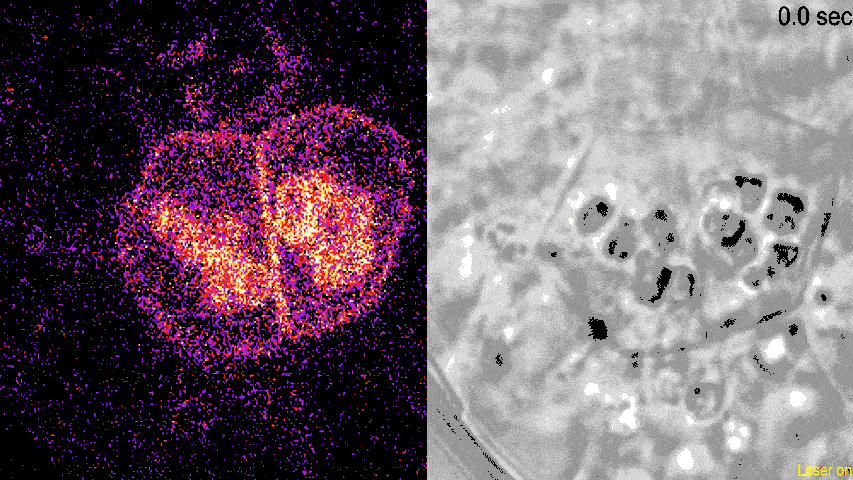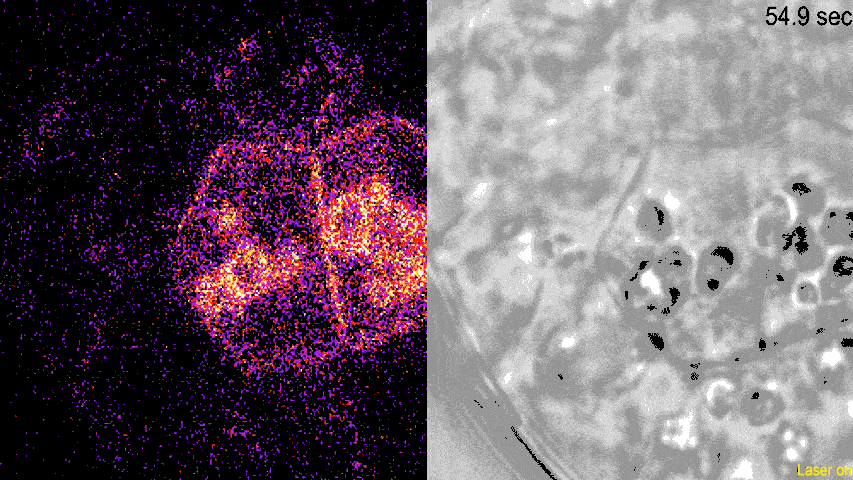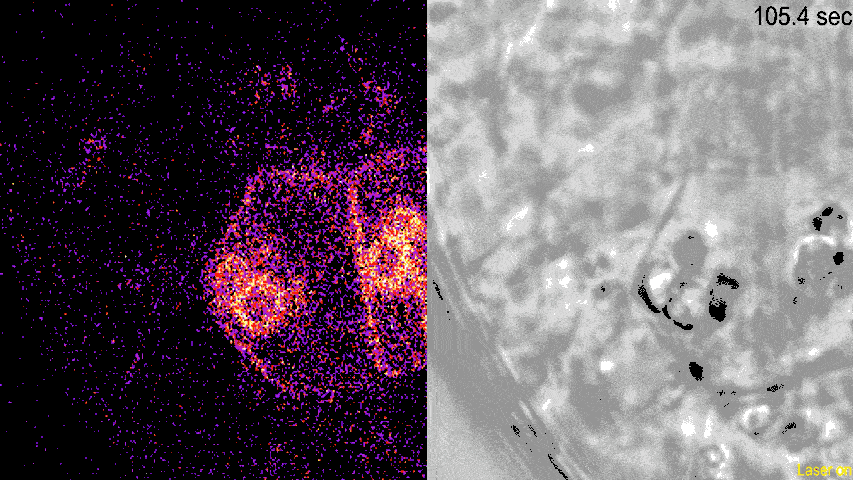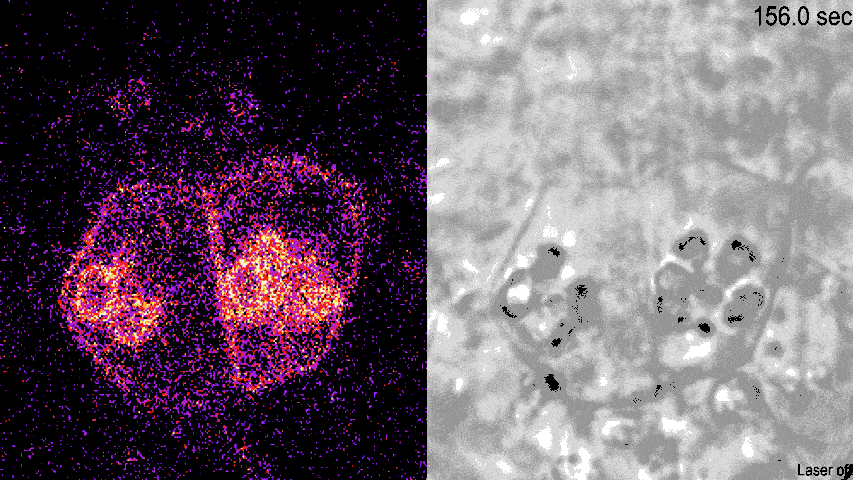
动图显示了荧光标记的LZY蛋白荧光(左)和明场下可见的淀粉体(右)。从荧光密度的变化可见,LZY蛋白对光学镊子操纵的淀粉体发生响应,进行相应的位置移动,并且向淀粉体集中的质膜位置聚集。
(图片来源:参考文献[4])
研究结果提出了一种重力传感机制,LZY蛋白通过传递淀粉体位置的信息来指示重力的方向。质膜上的LZY蛋白能够移动,当重力方向改变时,LZY蛋白可通过与淀粉体结合,重新定位到淀粉体沉积的位点,然后转移到质膜上新的部位产生极性。
研究还发现质膜上的LZY蛋白需要与细胞中一种称为RLD的蛋白结合,形成相互作用的分子伴侣,二者共同对生长素运输进行调节,很可能是将重力方向的信息通过膜转运的方式进行传递,从而影响生长素的运输和植物的生长变化。
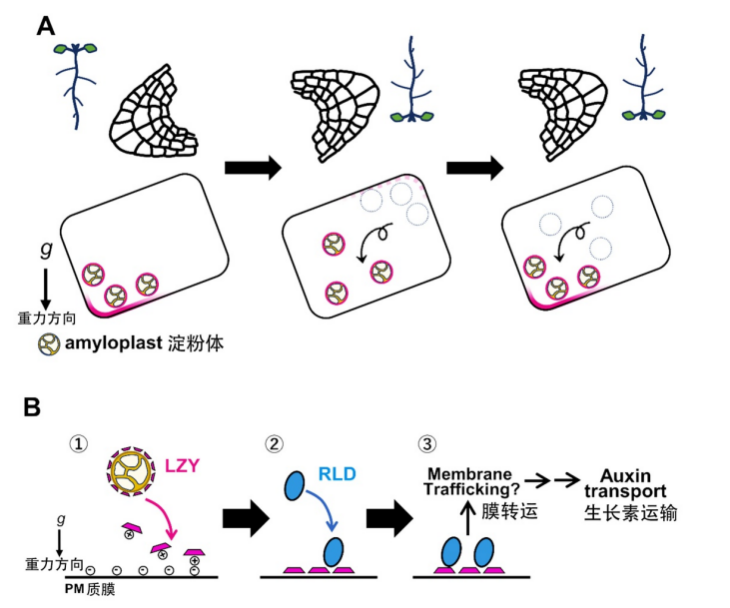
重力刺激下LZY蛋白依赖淀粉体沉积的亚细胞定位变化模型(A)和LZY蛋白在质膜上依赖淀粉体沉积的极性定位启动重力信号传导模型(B)
(图片来源:参考文献[4])
该发现的意义在于厘清了LZY蛋白在质膜的极性并不是通过重力实现,而是通过淀粉体的移动实现,也就是通过位置感应而不是通过机械力来感应的。
这说明,长期以来,植物对重力的感应被认为是“通过平衡石对细胞内部结构施加作用力实现”这一推测并不正确。这一发现有力地支持了位置传感器假说,为今后对植物极性运输的分子机制探索提供了极为重要的理论依据。
结语
科学家虽然弄清了植物是如何感知重力的,但是关于植物重力信号传递的分子机制,我们还有漫长的探索道路。其实,植物根系除了具有向地性,还具有向肥性、向水性等多种特性,根系会主动朝着含水丰富养分肥沃的土壤方向生长。正是植物根系的这些“聪明”特性,才让植物生命体得以很好地固着生长并受到保护,从而维系生长发育和繁殖的需求。
参考文献:
[1] Ajala C and Hasenstein K H. Plant Science, 2019, 285: 214–223.
[2] Kawamoto N and Morita M T. New Phytologist, 2022, 236: 1637–1654.
[3] Nakamura M, et al. Journal of Experimental Botany, 2019, 70(14): 3495–3506.
[4] Nishimura T, et al. Science, 2023,10.1126/science.adh9978.
[5] Singh M, et al. Frontiers in Plant Science, 2017, 8:1304.
[6] Stanga J, et al. Signaling in Plant Gravitropism. In: Signaling in Plants. Springer, Berlin, Heidelberg. 2009, pp209-237.
[7] Zhang Y, et al. New Phytologist, 2019, 224: 761–774.


 2023-11-11
2023-11-11


 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国