

信使RNA是由DNA的一条链作为模板转录而来的、携带遗传信息的能指导蛋白质合成的一类单链核糖核酸。
以细胞中基因为模板,依据碱基互补配对原则转录生成mRNA后,mRNA就含有与DNA分子中某些功能片段相对应的碱基序列,作为蛋白质生物合成的直接模板。mRNA虽然只占细胞总RNA的2%~5%,但种类最多,并且代谢十分活跃,是半衰期最短的一种RNA,合成后数分钟至数小时即被分解。1
简介信使核糖核酸信使RNA是指导蛋白质生物合成的直接模板。mRNA 占细胞内RNA总量的2%~ 5%,种类繁多,其分子大小差别非常大。2
信使RNA(mRNA)是一大类RNA分子,它将遗传信息从DNA传递到核糖体,在那里作为蛋白质合成模板并决定基因表达蛋白产物肽链的氨基酸序列。 RNA聚合酶将初级转录物mRNA(称为前mRNA)转录成加工过的成熟mRNA,这种成熟的mRNA被翻译成蛋白质。
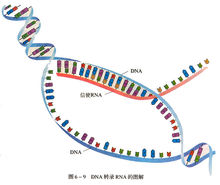 如在DNA中一样,mRNA遗传信息也保存在核苷酸序列中,其被排列成由每个三个碱基对组成的密码子。每个密码子编码特定氨基酸,但终止密码子例外,因为其终止蛋白质合成。
如在DNA中一样,mRNA遗传信息也保存在核苷酸序列中,其被排列成由每个三个碱基对组成的密码子。每个密码子编码特定氨基酸,但终止密码子例外,因为其终止蛋白质合成。
将密码子翻译成氨基酸的过程需要另外两种类型的RNA:转移RNA(tRNA)和核糖体RNA(rRNA)。tRNA介导密码子的识别并提供相应的氨基酸,rRNA是核糖体蛋白质制造机械的核心组成部分。
mRNA的存在首先由Jacques Monod和FrançoisJacob提出,随后由Jacob,Sydney Brenner和Matthew Meselson于1961年在加州理工学院发现。
原核生物和真核生物mRNA有不同的特点原核生物mRNA常以多顺反子的形式存在。真核生物mRNA一般以单顺反子的形式存在。3
原核生物mRNA的转录与翻译一般是偶联的,真核生物转录的mRNA前体则需经转录后加工,加工为成熟的mRNA与蛋白质结合生成信息体后才开始工作。
原核生物mRNA半寿期很短,一般为几分钟 ,最长只有数小时(RNA噬菌体中的RNA除外)。真核生物mRNA的半衰期较长, 如胚胎中的mRNA可达数日。
原核与真核生物mRNA的结构特点也不同。真核生物mRNA具有5‘帽子和3’多聚A尾巴,原核生物没有这样的首尾结构。
单顺反子与多顺反子mRNA翻译产物仅是单个蛋白质链(多肽)的mRNA称为单顺反子mRNA。大多数真核mRNA都属于单顺反子mRNA。多顺反子mRNA携带几个开放阅读框(ORF),每个开放阅读框都能被翻译成一条多肽,这些多肽通常具有相似的功能,通常构成最终复合蛋白的不同亚基。这些多肽链对应的DNA片断则位于同一转录单位内,享用同一对起点和终点。细菌和古细菌中的大多数mRNA是多顺反子的。
mRNA的环化在真核生物中,由于eIF4E和poly(A)结合蛋白之间的相互作用,mRNA分子形成环状结构,这两种结合蛋白都与eIF4G结合,形成mRNA-蛋白-mRNA桥。环化促进核糖体在mRNA上的循环,提高翻译效率,且确保仅有完整的mRNA得到翻译。
合成和加工mRNA分子的合成始于转录,并最终以降解结束。在被翻译之前,真核mRNA分子通常需要大量加工和转运,而原核mRNA分子则不需要。真核mRNA分子和它周围的蛋白质一起被称为信使RNP。
转录转录是指由DNA合成RNA的过程。在转录期间,RNA聚合酶根据需要将一个基因的DNA拷贝成mRNA,这个过程在真核生物和原核生物中是相似的。
与原核生物明显不同的是,真核RNA聚合酶在转录过程中与mRNA加工酶结合,因此,真核生物的mRNA加工可以在转录开始后快速进行。短寿命的未加工或部分加工的转录产品称为前体mRNA或pre-mRNA;一旦加工完全,它被称为成熟mRNA。
真核pre-mRNA加工mRNA的加工在真核生物、细菌和古细菌中差异很大。实质上,非真核mRNA在转录时是成熟的,除极少数情况外不需要加工。然而,真核pre-mRNA需要大量加工。
5’端加帽子:5‘ 帽(也称为RNA帽,RNA 7-甲基鸟苷帽或RNA m7G帽)就是一个经修饰的鸟嘌呤核苷酸,在转录开始不久后就被添加到新产生的真核mRNA的“前”即5'末端。 5’帽由末端7-甲基鸟苷残基组成,它通过5'-5'-三磷酸键与第一转录出的核苷酸连接。它的存在对于核糖体的识别和对mRNA的保护至关重要。
3’端加尾:是指聚腺苷酰基部分与mRNA分子的共价连接。在真核生物中,大多数信使RNA(mRNA)分子在3'末端被多聚腺苷酸化。Poly A尾巴和与其结合的蛋白质有助于保护mRNA免于被核酸外切酶降解。3’端加尾对于转录终止,从细胞核输出mRNA和翻译也很重要。 原核生物中的mRNA也常被3’端加尾,但此时的poly(A)尾巴促进而不是防止核酸外切酶对mRNA的降解。
mRNA的转运真核生物和原核生物之间的另一个区别是mRNA的转运。由于真核转录和翻译是在不同的细胞器内进行的,真核mRNA必须从细胞核输出到细胞质。 这一过程可能受不同信号通路的调节。成熟的mRNA通过其加工的修饰被识别,在结合帽结合蛋白CBP20和CBP80及转录/输出复合物(TREX)后通过核孔被输出到细胞质。
mRNA的翻译因为原核mRNA不需要加工或转运,所以原核生物mRNA在核糖体的翻译可以在转录结束后立即开始。因此,可以说原核生物的mRNA翻译与转录偶联发生。
已经加工并转运至细胞质的真核mRNA(即成熟mRNA)在核糖体的翻译发生在细胞质中自由漂浮的核糖体中,或者通过信号识别颗粒导向到的内质网中。因此,与原核生物不同,真核生物的mRNA翻译不直接与转录偶联。在某些情况下甚至可能发生这样的情况,即mRNA水平的降低却伴随着蛋白质水平的增加。
降解同一细胞内的不同mRNA具有不同的寿命(稳定性)。在细菌细胞中,单个mRNA可以存活数秒至超过一小时,但平均寿命为1至3分钟,因此,细菌mRNA的稳定性远低于真核mRNA。哺乳动物细胞mRNA的寿命从几分钟到几天不等。mRNA的稳定性越高,从该mRNA产生的蛋白质越多。 mRNA的有限寿命使细胞能够快速改变蛋白质合成以响应其不断变化的需求。有许多机制可导致mRNA的降解。
原核mRNA的降解原核生物mRNA的降解是不同核糖核酸酶包括核酸内切酶,3'核酸外切酶和5'核酸外切酶的共同作用的结果。在一些情况下,长度为数十至数百个核苷酸的小RNA分子(sRNA)可通过与互补序列碱基配对来促进RNase III对特定mRNA的降解。
真核mRNA的降解真核细胞的翻译和mRNA衰变之间存在着平衡。正在被翻译的mRNA被核糖体,真核起始因子eIF-4E和eIF-4G以及poly(A)结合蛋白结合,不能接触外泌体复合物,mRNA得到保护。mRNA的poly(A)尾巴被特异性外切核酸酶缩短,该核酸外切酶通过RNA上的顺式调节序列和反式作用RNA结合蛋白的组合定位到特定mRNA。 Poly(A)尾巴被去除破坏了mRNA的环状循环结构并降低了帽结合复合体的稳定性,导致mRNA会被外来体复合物或脱帽复合物降解。通过这种方式,可以快速降解翻译不活跃的mRNA,而翻译活跃的mRNA不受影响。
小干扰RNA在后生生物中,由Dicer产生的小干扰RNA(siRNA)被整合到称为RNA诱导沉默复合物(RISC)。该复合物含有内切核酸酶,切割与siRNA结合的完全互补的mRNA,产生的片段然后被核酸外切酶降解。 siRNA通常用于实验室细胞培养中阻断基因的功能。SiRNA被认为是病毒先天免疫系统的一部分,可以用于对双链RNA病毒的防御。
微小RNA微小RNA(miRNA)是小RNA,通常与后生生物mRNA中的序列部分互补。 miRNA与mRNA的结合可以抑制该mRNA的翻译并加速poly(A)尾部去除,从而加速mRNA降解。
提取、分离和纯化真核细胞的mRNA分子最显著的结构特征是具有5’端帽子结构(m7G)和3’端的Poly(A)尾巴。绝大多数哺乳类动物细胞mRNA的3’端存在20-30个腺苷酸组成的Poly(A)尾,通常用Poly(A+)表示。这种结构为真核mRNA的提取,提供了极为方便的选择性标志,寡聚(dT)纤维素或寡聚(U)琼脂糖亲合层析分离纯化mRNA的理论基础就在于此。mRNA的分离方法较多,其中以寡聚(dT)-纤维素柱层析法最为有效,已成为常规方法。此法利用mRNA 3’末端含有Poly(A+)的特点,在RNA流经寡聚(dT)纤维素柱时,在高盐缓冲液的作用下,mRNA被特异地结合在柱上,当逐渐降低盐的浓度时或在低盐溶液和蒸馏水的情况下,mRNA被洗脱,经过两次寡聚(dT)纤维柱后,即可得到较高纯度的mRNA。寡聚(dT)纤维素柱纯化mRNA。
本词条内容贡献者为:
张岩 - 副研究员 - 北京大学分子医学研究所

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国