

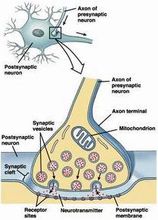
突触(synapse):两个神经元之间或神经元与效应器细胞之间相互接触、并借以传递信息的部位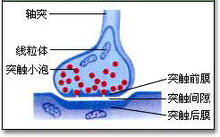 。突触一词首先由英国神经生理学家C.S.谢灵顿于1897年研究脊髓反射时引入生理学,用以表示中枢神经系统神经元之间相互接触并实 现功能联系的部位。而后,又被推广用来表示神经与效应器细胞间的功能关系部位。Synapse来自希腊语,原意是接触或接点。3
。突触一词首先由英国神经生理学家C.S.谢灵顿于1897年研究脊髓反射时引入生理学,用以表示中枢神经系统神经元之间相互接触并实 现功能联系的部位。而后,又被推广用来表示神经与效应器细胞间的功能关系部位。Synapse来自希腊语,原意是接触或接点。3
突触前细胞借助化学信号,即递质(见神经递质),将信息转送到突触后细胞者,称化学突触,借助于电信号传递信息者,称电突触。在哺乳动物进行突触传递的几乎都是化学突触;电突触主要见于鱼类和两栖类。根据突触前细胞传来的信号,是使突触后细胞的兴奋性上升或产生兴奋还是使其兴奋性下降或不易产生兴奋,化学和电突触都又相应地被分为兴奋性突触和抑制性突触。使下一个神经元产生兴奋的为兴奋性突触,对下一个神经元产生抑制效应的为抑制性突触。
化学突触或电突触均由突触前膜、突触后膜以及两膜间的窄缝——突触间隙所构成,但两者有着明显差异。胞体与胞体、树突与树突以及轴突与轴突之间都有突触形成,但常见的是某神经元的轴突与另一神经元的树突间所形成的轴突-树突突触,以及与胞体形成的轴突-胞体突触。
螯虾腹神经索中,外侧与运动巨大纤维间形成的突触便是兴奋性电突触。在螯虾螯肢开肌上既有兴奋性,也有抑制性化学突触。此外,尚发现一些同时是化学又是电的混合突触。
生物特点神经元之间神经冲动的传导是单方向传导,即神经冲动只能由一个神经元的轴突传导给另一个神经元的细胞体或树突,而不能向相反的方向传导。这是因为递质只在突触前神经元的轴突末梢释放。当神经冲动通过轴突传导到突触小体时,突触前膜对钙离子的通透性增加,突触间隙中的钙离子即进入突触小体内,促使突触小泡与突触前膜紧密融合,并出现破裂口。小泡内的递质释放到突触间隙中,并且经过弥散到达突触后膜,立即与突触后膜上的蛋白质受体结合,并且改变突触后膜对离子的通透性,引起突触后膜发生兴奋性或抑制性的变化。这里,递质起携带信息的作用。4
由于突触的单向传递,中枢神经系统内冲动的传递就有一定的方向,即由传入神经元传向中间神经元,再传向传出神经元,从而使整个神经系统的活动能够有规律地进行。中枢神经系统中任何反射活动,都需经过突触传递才能完成。
研究发展1896年C.S.Sherrington(C.S.谢灵顿)把神经元与神经元之间的机能接点命名为突触(synapse),当时他虽然还不了解接点的形态学,但是他指出神经元与神经元之间是不连续的,而且推论有些突触是兴奋![]() 性的有些突触是抑制性的。
性的有些突触是抑制性的。
在20世纪30、40年代对于突触之间是电学传递还是化学传递曾经发生过争论。现在知道有两类突触:电突触与化学突触。神经元之间化学传递的基本概念起源于哺乳动物内脏神经系统的研究。
20世纪初,J.N.Langley和他的学生发现肾上腺素的效应与刺激交感神经系统的效应十分相似。他的学生,T.R.Elliott甚至指出,肾上腺素可能是外周神经释放的化学刺激物。
后来H.H.Dale发现胆碱及其衍生物对心脏、膀胱和唾液腺的效应与刺激副交感神经相似,特别是乙酰胆碱最有效。Dale提出乙酰胆碱、肾上腺素的作用与刺激两类内脏神经的效应相似性的问题。
Otto Loewi在1921年所做的实验证明,刺激迷走神经释放活性化学物质,抑制心搏。继而证明,这种化学物质就是乙酰胆碱。
1936年Dale等人在刺激支配肌肉的运动神经后得到了神经释放的乙酰胆碱,因而把化学传递的假说推广到全部外周神经系统。证明乙酰胆碱是神经肌肉接点的神经递质后,直到1952年中枢神经系统的化学递质说才被广泛接受,而在7年之后,E.Furshpan和D.Potter又第一次清楚地证明了电突触的存在。
Furshpan和Potter在1959年首先指出在螯虾的可兴奋细胞之间有电学传递。电学传递可以发生在中枢神经系统的细胞之间、平滑肌细胞之间、心肌细胞之间、感受器细胞和感觉轴突之间。一个电突触的突触前膜和突触后膜紧紧贴在一起形成缝隙连接,电流经过缝隙连接从一个细胞很容易流到另一个细胞。向见图的A细胞送入阈下电脉冲,引起这个细胞膜电位的变化。如果送入细胞A的电流相当一部分经过缝隙连接流入细胞B,就会相应地在B细胞引起可以察觉的膜电位变化。由于只有一部分流入细胞A的电流进入细胞B,所以细胞B的膜电位变化比细胞A的小。
组成结构化学突触或电突触均由突触前、后膜以及两膜间的窄缝──突触间隙所构成(见图),但两者有着明显差异。胞体与胞体、树突与树突以及轴突与轴突之间都有突触形成,但常见的是某神经元的轴突与另一神经元的树突间所形成的轴突-树突突触,以及与胞体形成的轴突-胞体突触。
当轴突末梢与另一神经元的树突或胞体形成化学突触时,往往先形成膨大,称突触扣。扣内可见数量众多的直径在 30~150纳米的球形小泡,称突触泡,还有较多的线粒体。递质贮存于突触泡内。一般认为,直径为30~50纳米的电子透明小泡内贮存的是乙酰胆碱 (Ach)或氨基酸类递质。
有些突触扣内含有直径 80~150纳米的带芯突触泡和一些电子密度不同的较小突触泡,这些突触泡可能含有多肽。那些以生物胺为递质的突触内也含有不同电子密度的或大或小的突触泡。突触膜增厚也是化学突触的特点。高等动物中枢突触被分为GrayⅠ型和Ⅱ型,或简称Ⅰ型和Ⅱ型。前者的突触间隙宽约30纳米,后膜明显增厚,面积大;多见于轴突-树突突触;后者的突触间隙宽约20纳米,后膜只轻度增厚,面积小,多见于轴突-胞体突触。当然也存在介于两者之间的移行型。
电突触没有突触泡和线粒体的汇聚,它的两个突触膜曾一度被错误地认为是融合起来的,实际上两者之间有 2纳米的突触间隙;因此电突触又称间隙接头。电突触的两侧突触膜都无明显的增厚现象,膜内侧胞浆中也无突触泡的汇聚,但存在一些把两侧突触膜连接起来的、直径约2纳米的中空小桥,两侧神经元的胞浆(除大分子外)借以相通。如将化子量不大的荧光色素注入一侧胞浆中,往往可能过小桥孔扩散到另一神经元。这样的两个神经元,称色素耦联神经元。
化学突触的传递 冲动传到突触前末梢,触发前膜中的Ca离子通道开放,一定量的Ca离子顺浓度差流入突触扣。在Ca 离子的作用下一定数量的突触泡与突触前膜融合后开口,将内含的递质外排到突触间隙。此过程称胞吐。
被释放的递质,扩散通过突触间隙,到达突触后膜,与位于后膜中的受体结合,形成递质受体复合体,触发受体改变构型,开放通道,使某些特定离子得以沿各自浓度梯度流入或流出。这种离子流所携带的净电流,或使突触后膜出现去极化变化,称兴奋性突触后电位(EPSP),或使突触后膜出现超极化变化,称抑制性突触后电位(IPSP)。至今尚未发现兴奋性突触与抑制性突触在精细结构上的特征性区别,有人报道含圆形突触泡者为兴奋性突触,含椭圆形突触泡者为抑制性突触,但尚未得到进一步证实。
由细胞内记录的EPSP和IPSP都是迅速上升、缓慢下降、持续约30毫秒的局部电变化,只是在正常膜电位条件下前者为正,后者为负,以及IPSP的时程稍短些。
高等动物中枢每一突触后神经元上通常形成大量的突触(包括兴奋性和抑制性的),猫脊髓前角的一个运动神经元胞体上形成1200~1800个突触,约占据神经元胞体表面的38%。神经元通过对EPSP和IPSP进行空间总和(即对在神经元不同位置上出现的EPSP和IPSP进行总和)和时间总和(即对每个突触重复发生的突触后电位进行总和),以决定它产生兴奋还是抑制。总和后,如兴奋性突触后电位达到阈值,便触发动作电位。在突触传递中递质一旦释放,无论是否已与受体结合,便又迅速地被分解或被重吸收到突触扣内或扩散离开突触间隙,使突触得以为下次传递作好准备。
结构参数突触结构参数在CON组与LS组之间有明显不同,提示PNS已经引起子代海马突触发生形态学改变,可能对其可塑性有影响。突触可塑性是指突触在一定条件下调整功能、改变形态、增加或减少数目的能力,既包括传递效能的变化(LTP/LTD),也包括形态结构的变化,如PSD增厚或变薄等。一般认为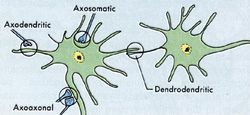 ,突触的修饰在很大程度上反映了整个神经系统回路的可塑性,因此也反映了行为的可塑性。
,突触的修饰在很大程度上反映了整个神经系统回路的可塑性,因此也反映了行为的可塑性。
LS组PSD厚度明显大于CON组(P

 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国